
Articles
The Living Soil
Oct
The Living Soil – Ethylene Oxygen Cycle
The Ethylene-Oxygen Cycle by Dr Alan Smith provides vital information and insights into how we care for the soil, our most precious resource. Just as relevent now as when it was first presented, this work is covered in the Living Soil module of my permaculture courses. It is republished here in response to requests from my students, and for the benefit of gardeners, farmers and all concerned with healthy soil and sustainable food production.
Smith’s conference paper, delivered shortly before his research was terminated in the late ’70s, was first published in one the early Permaculture Journals and republished in PIJ issue #39. I was editor at the time and wrote the forward to Smith’s paper. This is a very important soil process that explains the science of why Fukuoka’s no-till system was so successful and challenges many mainstream agriculture assumptions about the relationship between plants, soil microorganisms, pathogens and nutrient availability. – Robyn Francis
THE LIVING SOIL
No-dig gardeners and no-tillage farmers realise that healthy plants and good yields can be obtained on a sustained basis from undisturbed soils, nevertheless, for many lingering doubts remain. Doesn’t soil need to be periodically aerated to stimulate microbial activity and liberate nutrients to plants?
Soil scientist Alan Smith has been a principal research scientist for the New South Wales Department of Agriculture. PIJ put the question to him, his reply, and the article which follows contained some real surprises! “I can understand your confusion when trying to interpret the claims and counterclaims made regarding the value of ‘aerating’ soil. Obviously, I am a non-believer in the value of aeration, at least in Australian conditions. One thing that we must always be wary about are treatments that may give initial, short-term gains but lead to long-term problems. I believe the ‘aeration’ theory is such a treatment.
There is no doubt that ploughing soil does initially increase aeration and does result in intimate contact between the mineral soil and any organic residues. This stimulates microbial activity and nutrients immobilised in the organic reserves are liberated rapidly into the soil. However, unless plants are growing in the soil to take immediate advantage of these mobilised nutrients, they are leached or rapidly fixed in unavailable forms. ‘Aerating’ soil, of course, usually results in the removal of plant material and so there are no plants (or only a few) left to take advantage of the released nutrients. If this practice is continued season after season then it is obvious that a loss of nutrients results.
The ‘aeration theory’ really developed in the northern hemisphere where the extended cold winters prevent microbial decomposition of organic residues in soil. In spring it is advantageous to stimulate the decomposition rate so that plants can obtain nutrients during a relatively short growing season. In Australia, conditions are generally favourable right throughout winter for at least some organic matter breakdown. Thus, our conditions are very different. It is also worth considering just what problems arise when this ‘aeration’ is attempted in tropical soils where conditions are even more favourable for the breakdown of organic matter, Yes, we all recognise that under those conditions it is a recipe for disaster.”
Robyn Francis
 Ethylene Oxygen Cycle in Soil by Dr Alan Smith
Ethylene Oxygen Cycle in Soil by Dr Alan Smith
 Ethylene Oxygen Cycle in Soil by Dr Alan Smith
Ethylene Oxygen Cycle in Soil by Dr Alan SmithMICROBIAL INTERACTIONS IN SOIL AND HEALTHY PLANT GROWTH
Microbial interactions in soil play a key role in the biological control of plant diseases, the turnover of organic matter, and the recycling of essential plant nutrients. An understanding of the mechanisms involved may lead to more efficient methods of growing plants, whether they be food crops in agriculture or plants in gardens. Before these interactions can be discussed, however, it is essential to reaffirm the unique position that plants have in any ecosystem. They are the only living organisms that can directly utilise the energy of the sun and in the process they transform this energy into forms available to other living things. The green pigment, chlorophyll, in their leaves traps the light energy from the sun and an interaction occurs in leaves with carbon dioxide gas from the atmosphere to produce carbon compounds then available as energy sources to other living things, including man, other animals, insects and micro-organisms when they consume plants or plant remains.
Although plants have this unique ability to trap the energy of the sun and transform it into the chemical energy they need to grow, metabolise and reproduce, they also require other materials that they are unable to produce directly. For example, they require various elements, including nitrogen, phosphorus, sulphur, calcium, magnesium, potassium and trace elements. The soil is the reservoir of these elements, but to obtain adequate supplies plants must alter the environment around their roots to mobilise these nutrients. One important way the plant achieves this is by stimulating the activity of micro-organisms in soil around their roots and the microbes then enhance nutrient mobilisation. The plant stimulates microbial activity in soil by supplying chemical energy in the form of root exudates and litter. Thus, an intimate relationship exists between the plants and soil microbes.
Unfortunately, in many of the conventional methods used in agriculture this relationship is impaired, resulting in problems of nutrient supply to the plant and an increase in the incidence of disease. The latest research indicates that during the life of the plant up to 25 per cent of the chemical energy in the form of carbon compounds that is manufactured in the leaves is lost by the plant into the soil directly adjacent to the root. This material is lost either as root exudates or as dead, sloughed plant cells.
On a first examination this seems to be a highly inefficient, wasteful mechanism. The plant goes to considerable trouble to trap the energy of the sun and convert it to chemical energy, but then loses almost a quarter of the energy into the soil! One view is that nothing in nature is perfect and ‘leaky’ roots are inevitable. I certainly do not subscribe to this view. I firmly believe that if some living system is apparently wasting a quarter of the energy that is goes to the trouble to manufacture, then this loss must ultimately benefit the organisms directly. IF this is not the case, then evolution should have resulted in the selection of plants that lost less of their energy.
How does this loss of carbon compounds into the soil benefit the plant? Most importantly, these compounds are energy sources for the soil micro-organisms which proliferate in the rhizosphere, i.e. the soil zone directly adjacent to the plant root. These micro-organisms multiply so rapidly that they deplete the soil of oxygen at numerous microsites in the rhizosphere. Thus, oxygen-free anaerobic microsites are formed. The formation of these anaerobic microsites plays an important role in ensuring the health and vigor of plants.
ETHYLENE PRODUCTION IN SOIL
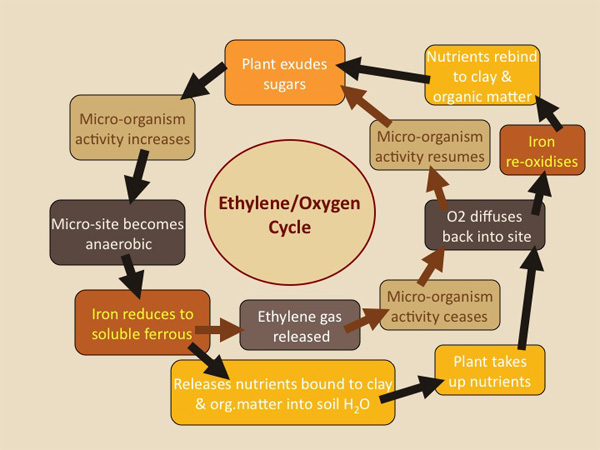
Our research shows that ethylene, a simple gaseous compound, is produced in these anaerobic microsites. Furthermore, this ethylene is a critical regulator of the activity of soil micro-organisms and, as such, affects the rate of turnover of organic matter, the recycling of plant nutrients and the incidence of soil-borne plant diseases. Concentrations of ethylene in the soil atmosphere rarely exceed 1 to 2 parts per million. Ethylene does not act by killing soil micro-organisms, but simply by temporarily inactivating them – when concentrations of ethylene in soil fall, microbial activity recommences.
Soil ethylene is produced in what we call the OXYGEN-ETHYLENE CYCLE.
Initially, the soil micro-organisms proliferate on the plant root exudates and deplete the soil of oxygen at microsites. Ethylene is then produced in these microsites and diffuses out, inactivating without killing the soil micro-organisms. When this occurs the demand for oxygen diminishes and oxygen diffuses back into the microsites. This stops or greatly reduces ethylene production, which enables the soil micro-organisms to recommence activity. Favourable conditions are then recreated for ethylene production and the cycle is continuously repeated. In undisturbed soils, such as found under forest and grasslands, ethylene can be continually detected in the soil atmosphere, indication that the oxygen-ethylene cycle is operation efficiently. Conversely, in most agricultural soils, ethylene concentrations are extremely low or non-existent. This is to be expected if ethylene plays an important role in regulation microbial activity in soil.
It is well established that in undisturbed ecosystems where there is a slow, balanced turnover of organic matter, efficient recycling of plant nutrients and soil-borne plant diseases are unimportant. When these ecosystems are disturbed for agricultural of forestry usage the situation changes dramatically, There is an alarming decline in the amount of soil organic matter, deficiencies of plant nutrients become commonplace and the incidence of plant disease increases dramatically.
We attempt to overcome these problems by additions of inorganic fertilisers and by the use of pesticides, which increase our production costs considerably. It is also generally true that the longer we farm soil, more and more of these inputs are necessary to maintain our yields. We argue that the trend could be reversed, at least partially, if we could create favourable conditions for ethylene production in these disturbed soils.
We now know that one of the major reasons why disturbed, agricultural soils fail to produce ethylene is because our techniques cause a change in the form of nitrogen in soil. In undisturbed soils, such as under forests or grasslands, virtually all the nitrogen present is in the ammonium form with just a trace of nitrate nitrogen present. When these ecosystems are disturbed for agricultural usage, virtually all the soil nitrogen occurs in the nitrate form. This change in form of nitrogen occurs because the disturbance associated with agricultural operations stimulates activity of a specific group of bacteria which convert ammonium nitrogen to nitrate nitrogen. Plants and micro-organisms can use either form of nitrogen, but our research has conclusively shown that ethylene production in soil in inhibited whenever the nitrate form is present at more than trace amounts.
Ammonium nitrogen has no such inhibitory effect on ethylene production. Nitrate nitrogen stops ethylene production because it interferes with the formation of the anaerobic microsites. When all the oxygen is consumed in the microsite a series of complex chemical changes then occur. One of the most important changes that occurs is that iron goes from the oxidised or ferric form to the reduced or ferrous form. Iron is one of the major constituents of soil, making up somewhere between 2 and 12% of its weight. In adequately aerated soil virtually all the iron exists as minute crystals of iron oxide and in this oxidised or ferric form is immobile in soil. If oxygen is completely consumed in microsites in soil, and reducing conditions exist, these minute crystals break down and iron is then transformed into the highly mobile ferrous or reduced form.
Again our research has shown that ethylene production occurs is soil only when iron is in the reduced or ferrous form. In other words, ferrous iron is a specific trigger for ethylene production. If there is no oxygen in the microsites, but nitrate nitrogen is present, then the complex chemical changes leading to the reduction of iron from the ferric to the ferrous form are inhibited. This is how nitrate nitrogen stops ethylene production.
How does ferrous iron trigger the release of soil ethylene? This form of iron reacts with a precursor of ethylene that is already present in the soil and a reaction occurs that results in the release of ethylene. Our work has established that this precursor originates from plants and, more importantly, it accumulates to appreciable amounts only in old, senescent plant leaves. When these old leaves fall to the ground and decompose, the precursor accumulates the soil. Then, when conditions become favourable for mobilisation of ferrous iron, ethylene is produced.
We have also shown that different plant species vary markedly in the quantities of precursor that accumulate in their old leaves. This is important to know when selection plant species to use as cover crops to increase the ability of agricultural soils to produce ethylene. A few of the plant species that produce high concentrations of precursor are rice, phalaris, chrysanthemum, avocado, bullrush and Pinus radiata. Some of the low producers include Dolichos, paspalum, lucerne and bracken fern. In retrospect it should not be too surprising that the ethylene precursor accumulates appreciably only in old, dead plant leaves. After all, in natural communities of plants old dead leaves comprise the bulk of the litter that falls on to soil. Also, it is equally clear that in agricultural situation most of the old plant leaves are removed either during harvest or by grazing or by burning crop residues. Thus, agricultural soils are usually deficient in precursor.
It is now possible to specify the soil conditions necessary for ethylene production –
(1) there must initially be intense aerobic microbial activity, at least in the rhizosphere, to ensure that oxygen-free, anaerobic microsites form;
(2) conditions in the microsites must become sufficiently reduced to mobilise ferrous iron to trigger ethylene release;
(3) concentrations of nitrate nitrogen in soil must be kept to trace amounts, otherwise ferrous iron will not be mobilised;
(4) there must be adequate reserves of the ethylene precursor in soil.
MOBILISATION OF ESSENTIAL PLANT NUTRIENTS
A major limitation to plant growth in most agricultural soils is an inadequate supply of essential plant nutrients. This occurs even though there are adequate reserves of these nutrients in soil, but they are held in highly insoluble forms. Their high degree of insolubility prevents loss from the soil by leaching, but since they are only available to the plant in the soluble form, problems of supply rate to plants are created. Formation of anaerobic microsites in the rhizosphere of plants, which is of such paramount importance to ethylene production, can play a critical role in the mobilisation and thus supply rate of these essential nutrients to plants. This mechanism revolves around the importance of iron in soil.
As already discussed, under normal conditions in soil most of the iron occurs as minute crystals of iron oxide. These crystals have a large surface area and are highly charged. As a result plant nutrients such as phosphate, sulphate and trace elements are tightly bound to the surfaces of these crystals. In this form they are virtually unavailable to plants. If, however, anaerobic microsites develop, these crystals break down and the bound nutrients are released for uptake by the plant. At the same time high concentrations of ferrous (reduced and mobile form) iron are released into the soil solution in the microsite. The other essential plant nutrients, including calcium, potassium, magnesium and ammonium, are held on the surfaces of clay and organic matter. When concentrations of ferrous iron increase so much, these nutrients are displaced by the ferrous iron into the soil solution, where they too are now available for uptake by plant roots.
Since anaerobic microsites are most likely to form in the rhizosphere of plants, the nutrients are mobilised exactly where they are required by the plant. An additional advantage of this mechanism is that if the released nutrients are not utilised by plant roots they cannot be leached in the soil. As soon as they migrate to the edge of the anaerobic microsite, reoxidation of the iron occurs with recrystallisation of iron oxide. These crystals then rebind the nutrients and prevent their loss by leaching.
The soil conditions necessary for this mechanism to operate are identical with those required for ethylene production. Thus in agricultural soils, where ethylene production is inhibited or impaired, this mechanism of nutrient mobilisation is also restricted. Again, under these conditions, the elevated concentrations of nitrate nitrogen that occur in agricultural soils are a major inhibitor of efficient nutrient mobilisation.
Successful management of soils to increase the likeihood of anaerobic microsite formation, which will help ensure a balanced oxygen-ethylene cycle and enhance mobilisation of essential plant nutrients, will demand alterations to some of the established practices in agriculture. For example, techniques aimed at increasing aeration and the oxidation states of soil, which give short-term increases in plant growth but rapidly create lone-term problems of nutrient depletion and increased plant disease incidence, will require modification.
Treatments which stimulate rates of nitrification (transformation of ammonium nitrogen to nitrate nitrogen), such as excessive use of nitrogenous fertilisers, overuse of legume dominant pastures, or excessive removal of plants by overgrazing or forestry operations, require re-examination.
Some practical guidelines for successful management of soils include:-
(1) It is essential that organic residues be returned continually to the soil. Organic residues contain essential plant nutrients for recycling, stimulate microbial activity in soil, supply ethylene precursor, and restrict the rate of nitrification in soil. It is best to use mature plants as a source of organic amendments and it is better to return the residues to the soil surface rather than incorporate the into the soil.
(2) Techniques of minimum tillage should be utilised wherever practical. This ensures that plants are growing in soil virtually continually, that there is a minimum of disturbance to the soil and increases the amount of organic matter that is returned to the soil. Again, nitrification is restricted when these techniques are used.
(3) Whenever soil is amended with nitrogenous fertiliser it is best to apply the nitrogen in the ammonium form and to apply it in several small applications rather than one or two heavy dressings. This again restricts the chance of nitrification.
(4) In some situations it will be advisable to add chemical inhibitors of nitrification (e.g. N-Serve or Terrazole) to soil with the nitrogenous amendments to further ensure that nitrification is restricted.
Alan Smith, 1977
This article first appeared in ‘Australian Plants’ Vol. 9 No. 73, 1977, then was published in issue #7 of the International Permaculture Journal in March 1981 and was republished in the PIJ #39 in 1992.
Diagram and photo by Robyn Francus, Permaculture Education